| lab a-z index | phone book |
| June 22, 2005 | news releases | receive our news releases by email | science@berkeley lab |
|
|||
|
|||
| The Making and Breaking of Microtubules
How a Nucleotide Controls the Flexibility of Tubulin |
||||||||||||||||||||||||||||||||||||||||
| Contact: Paul Preuss, (510) 486-6249, paul_preuss@lbl.gov | ||||||||||||||||||||||||||||||||||||||||
| BERKELEY, CA – Microtubules are active protein polymers critical to the structure and function of cells and the process of cell division. In a living cell their growing ends constantly elongate and retreat in a thrashing frenzy of polymerization and depolymerization, like the writhing snakes of Medusa's hair. Known prosaically as "dynamic instability," this ongoing rapid growth and shrinkage is key to the diverse workings of microtubules in the cell.
For the first time, researchers have revealed at molecular level the forms taken by transitional structures of tubulin (the protein from which microtubules are formed) during the assembly and disassembly of microtubules. Details of these peculiar structures show how the binding to tubulin of the nucleotide guanosine triphosphate, GTP, controls activity at the growing end of the microtubule. Eva Nogales and Hong-Wei Wang of the Life Sciences Division of the Department of Energy's Lawrence Berkeley National Laboratory report their findings in the 16 June 2005, issue of the journal Nature. "Tubulin is a major target for anticancer drugs, which can prevent the transition from growing to shrinking states or vice versa," says Nogales, who is also a member of Berkeley Lab's Physical Biosciences Division, an associate professor of biochemistry and molecular biology at the University of California at Berkeley, and a Howard Hughes Medical Institute Investigator. "We had previously produced high-resolution structures of stabilized tubulin structures, but nobody knew the structure of the transitional states. Our goal was to understand the process of microtubule assembly and disassembly." Says Wang, "Our current work actually revealed the structures of different tubulin assemblies at the very ends of growing microtubules and depolymerizing microtubules. This allows a new understanding of the mechanism of microtubule dynamic instability, opening opportunities for new experiments testing the physiological roles of these microtubule intermediates in the cell." Richard Rodewald, a program officer and cell biologist at the National Institute of General Medical Sciences, which funded the research, noted that "these studies provide a persuasive model, in elegant atomic detail, for how binding of GTP to a specific site in tubulin leads to far-reaching structural changes that drive microtubule growth." Building a microtubuleMicrotubules are polymers whose basic units are pairs (dimers) of similar but not identical tubulin proteins, dubbed the alpha and beta forms. During polymerization the dimers stack end to end to make a protofilament. About thirteen protofilaments are arranged side by side, extending longitudinally, to form the walls of a cylindrical microtubule.
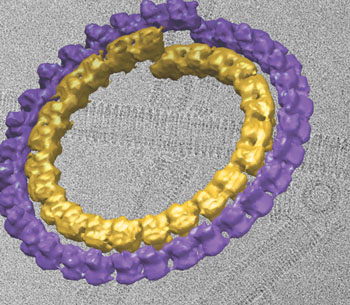
The so-called minus end of the microtubule grows slowly and is often anchored to a cellular structure. The other end, the plus end, is a hotbed of activity. In the presence of GTP the microtubule's protofilaments acquire more tubulin dimers, and the whole microtubule extends rapidly for many millionths of a meter before suddenly switching off and shrinking again. Nucleotides like GTP are best known as units of the nucleic acids DNA and RNA, consisting of a base (a letter in the genetic code) plus a sugar and three phosphate groups. But these molecules also regulate enzyme activity and play a crucial role in controlling microtubule growth. When GTP binds to a particular locus on beta tubulin (the E-site or exchangeable site) polymerization occurs: tubulin dimers pile onto the protofilaments, and the microtubule grows. But through hydrolysis (the word means "water splitting"), GTP readily changes to a different nucleotide, guanosine diphosphate, or GDP. GDP-bound tubulin depolymerizes the microtubule by curling up protofilaments at its plus end and pulling it apart. Freezing the frenzyAlthough tubulin was known to polymerize in the presence of GTP and depolymerize in the presence of GDP, before Nogales and Wang it was impossible to visualize what was happening structurally during these rapid transitional states. Then, says Nogales, "Hong-Wei found biochemical tricks for slowing down the transitions almost to the point of stopping them." Cryo-electron microscopy (cryo-EM) is capable of capturing such polymer structures without having to break them down and crystallize single subunits for x-ray diffraction analysis. X‑ray diffraction might allow higher resolution, but it would lose all information concerning contacts between the subunits during polymerization. "With cryo-EM, by changing the parameters we could see how different polymer forms convert," says Nogales. Nogales and Wang first tackled the depolymerization process, watching what happens to microtubules when GDP is bound to tubulin. During this process tubulin protofilaments curve back sharply from the end of the microtubule, forming "peels," like a wood shaving curling up from a plane. Wang found conditions for which the curls of GDP-bound tubulin close into tubes, with the protofilaments tightly wrapped around the tube axis. These tubes are double-layered, a property that contributes to their stability — but it also put the brakes on the project for a long time.
"The key was to get the tubulin assemblies locked at the correct states suitable for structural analysis, but even more challenging was finding an algorithm to reconstruct the double-layered GDP-tubulin tubes," says Wang. "Due to the double-layered property, it was impossible for us to use classical helical reconstruction methods," he says, "but there were no other methods available when we started the project. We had to spend over two years developing and implementing our own algorithm. Once it was ready, we were able to get the correct reconstruction in less than three months." Next the researchers tackled the polymerization process, watching what happens when GTP is bound to tubulin molecules. If GDP somehow causes protofilaments to curl up tightly, GTP does the opposite, causing protofilaments to straighten and assemble themselves into microtubules. But because GTP can rapidly hydrolyze into GDP, capturing transitional states of GTP-bound tubulin directly was impractical. Instead the researchers worked with a similar molecule, GMPCPP, an analogue of GTP that binds to the same site but is not vulnerable to hydrolysis. Using cryo-EM they were able to capture GMPCPP-bound tubulin during the initial steps of microtubule growth.
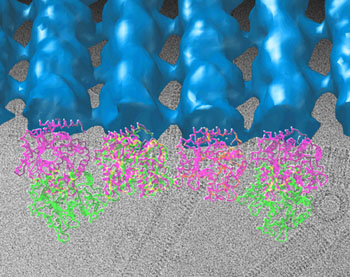
At low temperatures, GMPCPP-bound tubulin forms curved ribbons of a few protofilaments side by side; as they grow wider the ribbons close into very large helical tubes with a diameter of 500 angstroms, instead of the slim, 25-angstrom diameter of a microtubule (an angstrom is a ten-billionth of a meter, roughly the dimension of a small atom). Raised to body temperature, however, the protofilamentary ribbons immediately straighten and convert to normal microtubules, suggesting that the ribbons and gently curved sheets correspond to the polymerizing protofilaments at the end of a growing microtubule. Making contactsIn 1998, when she was a postdoctoral fellow in the laboratory of Kenneth Downing, a senior scientist in Berkeley Lab's Life Sciences Division, Nogales obtained the atomic structure of tubulin in its stable, straight form, using electron crystallography methods, in which two-dimensional arrays of tubulin "flat" polymers were studied using cryo-EM and electron diffraction. By "docking" this crystallographic model within the structures they have now obtained for the polymerization intermediates at more modest resolution, Nogales and Wang have been able to see changes in tubulin molecules, just how their capacity to interact with other molecules is altered by binding to GTP or GDP, and how these alterations control bending and flexibility. They found that in GDP-bound tubulin, the contacts between the alpha and beta tubulins within and between dimers are both affected, although in significantly different ways, resulting in a curved protofilament that cannot form lateral contacts. By contrast, in tubulin that binds the GTP analogue GMPCPP, contacts between the subunits are straight enough to allow lateral interaction between growing protofilaments. Says Nogales, "Such contacts appear semiconserved with respect of those previously seen in microtubules, and suggest a mechanism by which microtubules will first grow into open sheets that are able to quickly 'zip' into a close tube." Interestingly, in both polymerizing and depolymerizing transitional states, protofilaments usually occur in pairs. This is puzzling, because the most common microtubule has 13 protofilaments, which points to a change in the number of protofilaments after GTP hydrolysis, by a mechanism not yet understood. The new high-resolution models of tubulin transitional states will be used to understand how microtubules explore their cellular environment to find their goals — a process crucial to the accurate deployment of spindles during cell division, for example — and how drugs can be designed and targeted to put a monkey wrench into the growth of cancer cells. "Nucleotide-dependent bending flexibility of tubulin regulates microtubule assembly," by Hong-Wei Wang and Eva Nogales, appears in the 16 June 2005 issue of Nature. This research was funded by the National Institute of General Medical Sciences in the National Institutes of Health. Eva Nogales's laboratory is also supported by the Department of Energy and the Howard Hughes Medical Institute. Berkeley Lab is a U.S. Department of Energy national laboratory located in Berkeley, California. It conducts unclassified scientific research and is managed by the University of California. Visit our website at http://www.lbl.gov. |
||||||||||||||||||||||||||||||||||||||||
| Top | ||||||||||||||||||||||||||||||||||||||||