|
November 15, 2002
|
||
|
 |
 |
| How a structural protein regulates genes in immune-system cells, part 2 | ||
| Contact: Paul Preuss, paul_preuss@lbl.gov | ||
|
|
Down for the count
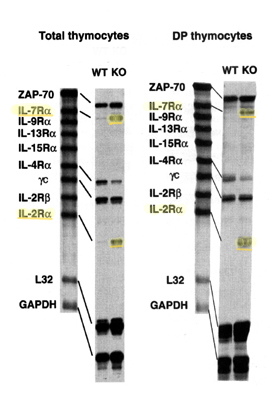
Kohwi-Shigematsu and her colleagues pinpointed SATB1's vital role in T-cell development by preparing "knockout" mice that were genetically normal in all respects except that they lacked the gene for SATB1, and thus the SATB1 protein itself. Designated SATB1-null, the knockout mice had problems from the start. Some of their reflexive behaviors were odd. Although most of their organs were normal, their thymus, spleen, and lymph nodes were smaller than those of the normal "wild" type. They were small and thin and lived only three weeks; wild-type mice live two years or more. Physiological causes soon emerged: lack of SATB1 had wrecked the immune system of the knockout mice. Very few immature thymocytes were produced to begin with, and those that progressed to the crucial double-positive stage inappropriately expressed receptors and markers. It was at this stage that most T-cell development in the SATB1-null mice came to a halt; while a very low number of helper T cells were produced, killer T cells were absent. Thymocytes that did develop to the double-positive stage expressed proteins, like interleukin-2 receptors, that should normally be repressed at this stage. Worse, the signals that activate normal mature T cells, inducing them to produce interleukin-2 and other proteins, instead caused the few helper T cells from SATB1 knockout mice to die off through apoptosis, programmed cell suicide.
Numerous genes essential to T-cell development and function were unregulated or misregulated, including genes for interleukin receptors, a gene for a chemokine receptors that stimulates activity in immune-system cells, genes that code for DNA binding proteins, genes for cell-surface markers, and genes related to apoptosis and cell proliferation. Nor is expression of SATB1 restricted to thymocytes: it is also found in other specific precursor/progenitor cells, including osteoblast cells that become bone. Runaway apoptosis of cells that normally express SATB1 is what may have killed the knockout mice at the end of their short lives. "In the absence of SATB1, hundreds of genes are dysregulated in thymocytes," says Kohwi-Shigematsu. "SATB1 tells the cells when and which genes should be expressed or remain silent." In thymocytes, SATB1 appears to regulate up to two percent of all the genes in the mouse genome. An architect of the chromatin Evidence for what began as a hypothesis by Kohwi-Shigematsu and her colleagues was fast accumulating: SATB1 forms a physical network inside the nucleus of thymocytes, one that resists salt extraction and the removal of genomic DNA and RNA. The SATB1 network actively tethers specialized DNA sequences and organizes the chromosomes into distinct loop domains, containing specific sets of genes and other DNA. SATB1 is not just a chromosome organizer, however. It orchestrates gene expression by allowing chromatin remodeling to take place at specific genomic locations. To find the proteins that associated with SATB1 at specific gene locations, Kohwi-Shigematsu and her colleagues applied techniques with arcane names like DNA-affinity chromatography and chromatin immunoprecipitation. What they found were protein complexes known to be involved in chromatin remodeling and gene regulation. These included ACF1, "ATP-utilizing chromatin assembly and remodeling factor 1," and ISWI, "imitation switch," named for its resemblance to yeast's SWI switch, a component of one of the first-identified molecular machines for remodeling chromatin. ACF1 and ISWI are members of CHRAC, the "chromosome accessibility complex," which shuttles nucleosome beads back and forth like an accountant manipulating an abacus. Another complex, the "nucleosome remodeling and histone deacetylation" (NuRD) complex, works directly on the histone units at the cores of nucleosome beads. Histone proteins, rather like New World monkeys, have prehensile "tails" with which they bind themselves together, eight to a nucleosome spool, or bind the DNA to the spool, or bind the nucleosome to other nucleosomes and different kinds of histones. Histone tails grasp tightly if deacetylated (lacking a particular group of carbon, hydrogen, and oxygen atoms); when acetylated, they let go — a necessary step in allowing DNA to be read by enzymes for gene transcription.
The researchers have shown, explicitly in the case of the interleukin-2 receptor-alpha gene (IL-2Ra) and in other genes as well, that SATB1 controls gene expression — in some cases up to tens of thousands of bases away from the attachment site — by providing docking sites for these remodeling complexes, which alter the shape of nucleosome arrays and activate and deactivate DNA transcription. In microscope images collected with the aid of Life Sciences researcher Abby Dernburg, the network in the cell nucleus stands out clearly as a cage-like structure of fluorescently stained SATB1 proteins circumscribing heterochromatin. (Heterochromatin is a densely packed type of chromatin associated with silent genes.) The cage-like SATB1 structure provides the sites where the bases of the looped chromatin meet chromatin-remodeling and modifying factors, for which it functions as a platform. How does SATB1 know whether to activate or repress transcription of a gene at a given time? Whether chromatin remodeling is motivated by demand from below or orders from above, Kohwi-Shigematsu says, "is a chicken and egg question" — one that will keep her and her colleagues busy for the foreseeable future, in the search for even deeper secrets of gene regulation. Additional information
|
||||||||||||||||||||||||||||