|
March 29, 2002
|
|
 |
 |
| New Cancer Role Found for Cell-Dividing Protein | ||
| Contact: Lynn Yarris, lcyarris@lbl.gov | ||
|
|
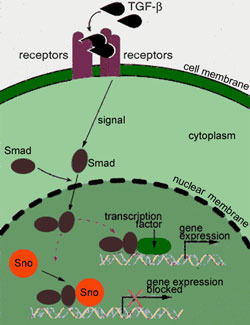
A protein long known to play a vital role in the process by which a living cell divides into two new "daughter" cells has now been shown to also play an important role in cancer. Researchers with Lawrence Berkeley National Laboratory (Berkeley Lab) have shown that the anaphase-promoting complex (APC) of proteins that breaks apart the connections between daughter cells during cell division also breaks apart a protein complex within the cell nucleus that is important for maintaining normal cell growth. "Until now, the only role known for APC is that it unglues the chromatids, or chromosome strands, to allow the separation of two daughter cells during mitosis," says Kunxin Luo, a cell biologist with Berkeley Lab's Life Sciences Division, who led this research. "Our studies reveal a novel role for APC in the regulation of TGF-beta signaling as well." TGF-beta, which stands for transforming growth factor-beta, is an extracellular protein responsible for controlling the growth and differentiation of epithelial cells—the cells that line the skin, kidney, glands, lungs, gastrointestinal tract, bladder, and blood vessels. The TGF-beta signal activates genes that, among other things, instruct an epithelial cell to stop growing. If something goes awry, causing the TGF-beta signal to be blocked, cell growth can continue unchecked, giving rise to cancerous tumors. Nearly 90 percent of all human cancers involve epithelial cells. Luo, who holds a joint appointment as an assistant adjunct professor with the University of California at Berkeley (UCB), led a previous study a year ago which reported that the TGF-beta signal can be blocked by "Ski" and "Sno," two closely-related proteins, long-suspected of being major contributors to the development of a number of cancers. Luo's research group demonstrated that the Ski and Sno oncoproteins interact with a family of tumor-suppressing proteins called "Smad" to regulate the TGF-beta signal. "Our work suggested that cancer development is a delicate balancing act in which the balance gets tipped the wrong way," Luo said at the time. To this protein balancing act, she and her colleagues have now added APC—and in the process have uncovered new evidence that the signals regulating vital cell processes arise from a highly sophisticated network of interacting proteins. "We've now shown that APC is a key component in the Smad-induced
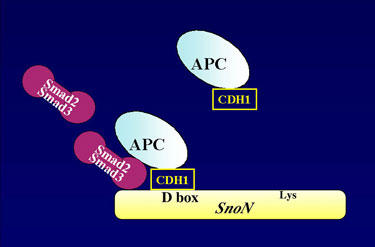
degradation of Sno protein," Luo says, "and this mechanism can
help provide insights into what causes a normal cell to become cancerous."
TGF-beta proteins cannot themselves enter a living cell and must therefore transmit their signals by attaching themselves to receptor proteins on a cell's outer surface. The signal generated by this interaction is then ferried across the cell membrane, through the cytoplasm, and into the nucleus via Smad proteins. Luo and her colleagues had previously shown that a normal level of Sno inside the nucleus blunts TGF-beta signals, but as the number of Smad proteins increases, the level of Sno degrades until it is low enough for the signals to take effect. With the results of this latest study, they now understand the mechanism by which the degradation of SnoN, the most predominant form of the Sno protein, takes place. "When the Smad proteins enter the nucleus they pick up APC proteins and bring them along to chop up the SnoN," explains Luo. "The SnoN contains a region called a destruction box that the APC targets. This is a similar molecular motif to what the APC targets in cyclins and other cell cycle-regulated proteins."
With the levels of SnoN reduced, TGF-beta signals get through and their target genes become activated. After about two hours, the TGF-beta signal has resulted in a marked increase in the expression of the Sno gene, which causes the level of SnoN to once again rise until there's enough present to block the signal and allow cell growth to resume. The mechanism is a negative feedback loop that must be maintained for normal growth activity. The next step, Luo says, is to determine whether high levels of Ski and Sno proteins alone are enough to cause a normal epithelial cell to become cancerous. It is known that mice cannot survive long if the Ski and Sno genes are entirely removed, so she and her colleagues have developed a mouse that carries mutant forms of the genes, which produce proteins that cannot block the TGF-beta signal. They anticipate results later this year. Members of Luo's team included Shannon Stroschein, who is with Berkeley
Lab and UC Berkeley, and Shirin Bonni and Jeffrey Wrana, from the Samuel
Lunenfield Research Institute in Toronto. Additional information:
|
||||||||||||||||||||||||