| lab a-z index | phone book |
| November 8, 2005 | news releases | receive our news releases by email | science@berkeley lab |
|
|||
|
|||
Discovering the First Steps in Transcription-Coupled RepairAn Important DNA Repair Mechanism Linked to Premature Aging Yields |
||||||||||||||||
| Scientific Contact:
Priscilla Cooper, (510) 486-7346, pkcooper@lbl.gov Media Contact: Paul Preuss, (510) 486-6249, paul_preuss@lbl.gov |
||||||||||||||||
| BERKELEY, CA – A team of scientists led by Priscilla Cooper, a senior staff scientist in the Life Sciences Division of the Department of Energy's Lawrence Berkeley National Laboratory, has discovered new players in the first steps of transcription-coupled repair (TCR), an essential but still mysterious mechanism of DNA repair.
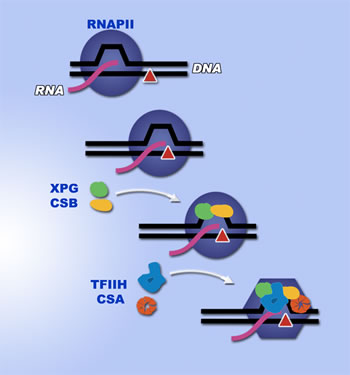
If a blockage occurs when genetic information from a cell's DNA is being transcribed into RNA — an activity vital to the synthesis of proteins — the transcription-coupled repair (TCR) process detects the obstruction and repairs the damage. But when TCR itself fails, the results can be lethal to cells and to the organism. In humans, failure of transcription-coupled repair is the cause of Cockayne Syndrome, an extreme form of accelerated aging that is inevitably fatal early in life. Cooper and her colleagues illuminated, for the first time, the roles played by two important proteins in recognizing blockages in transcription and in initiating an efficient method of repair; their results suggested a previously unsuspected mechanism for the repair process. The team, whose members hold positions at Berkeley Lab, the University of California at Berkeley, the Howard Hughes Medical Institute, the Skaggs Institute for Chemical Biology, and the Scripps Research Institute, includes Altaf Sarker, Susan Tsutakawa, Seth Kostek, Cliff Ng, David Shin, Marian Peris, Eric Campeau, John Tainer, and Eva Nogales, in addition to Cooper. Their results appear in the October 28, 2005 issue of the journal Molecular Cell. DNA repair: keeping up with lifeDNA is constantly under attack from sources inside and outside the body, including sunlight, ionizing radiation, other environmental carcinogens, and free radicals from the cellular metabolism. DNA damage ranges from one or a few altered nucleotides in a single strand of the double helix, to breaks in one or both strands and crosslinks between the two strands. To prevent accumulation of mutations and the production of altered proteins, cells deploy an arsenal of repair mechanisms to excise and replace defective nucleotides, reconnect broken strands, and patch up other kinds of damage. Transcription-coupled repair is unique: it targets repair to genes that are actively being transcribed into messenger RNA. TCR was discovered about 20 years ago as a result of comparing the properties of cells from patients with Cockayne Syndrome (CS) versus those with xeroderma pigmentosum (XP), two different hereditary diseases with a common feature: both entail extreme sensitivity to sunlight. However, in XP there is loss of ability to repair damage to DNA caused by ultraviolet radiation throughout the genome, whereas in CS the global repair mechanism is intact but transcription-coupled repair is defective. Thus active genes cannot be repaired preferentially. In XP patients, exposure to sunlight typically causes hyper-pigmented skin that is dry and parchment-like, and is followed by multiple skin cancers. If carefully shielded from ultraviolet light, for example by window filters and protective clothing, many XP sufferers can lead seemingly normal lives. XP results from mutations in any one of seven genes, labeled XPA through XPG, which are involved in the well-understood DNA repair mechanism called nucleotide-excision repair. In contrast, Cockayne Syndrome is marked not by skin cancer but by severe physical and mental retardation — victims have an unusually small brain and fail to grow and develop normally after birth; pronounced wasting usually begins in the first year of life. As they grow older, CS sufferers look increasingly aged, with faces marked by sunken eyes. Average life expectancy is only 12 years and few survive their teens. Cockayne Syndrome usually results from mutations in one of two genes, CSA or CSB, although mutations in three XP-associated genes — XPB, XPD, and XPG — can also cause clinical CS. When transcription failsGenes are copied into messenger RNA by the large, complex molecular machine known as RNA polymerase II (RNAPII). RNAPII works by separating the two strands of the DNA helix — opening what's called a "transcription bubble" — and moving along the strand to be transcribed like an engine along a cog railway, copying the genetic information in the gene to create a length of messenger RNA as it goes. As the RNAPII engine trundles along, enclosing the DNA and the open bubble, it sometimes encounters a damaged nucleotide on the strand it is transcribing — the sort of damage that might have been inflicted by ultraviolet radiation or oxidative damage, for example. Or it may encounter situations such as a highly repetitive sequence of bases, with folding of the DNA upon itself, which make progress difficult. Unable to continue copying, RNAPII pauses and sometimes grinds to a halt. Exposure to sunlight causes decreased transcription when ultraviolet-induced DNA damage blocks RNAPII. Transcription-coupled repair normally removes the blocking damage and allows transcription to recover; in Cockayne Syndrome, however, transcription cannot resume, and the affected cells die. "It has been assumed that defects in transcription-coupled repair are the cause of Cockayne Syndrome, although the question remained whether was the problem was inability to deal with damaged DNA or a subtle defect in general transcription," says Cooper. "To understand the exact nature of the problem we first needed to know how stalled RNAPII is recognized." The protein CSB, which in a mutated form causes one kind of Cockayne Syndrome, was known to recognize stalled RNAPII; CSB was presumed to be involved in initiating transcription-coupled repair, but by what means was unknown. In addition, there were hints for an important but unknown role for the protein XPG in transcription-coupled repair. In both humans and mice, when gene mutations produce a severely truncated form of XPG, the result is profound Cockayne Syndrome and very early death. Through an ingenious series of biochemical experiments, Cooper and her colleagues discovered evidence that XPG and CSB must work together in recognizing stalled RNAPII. The researchers learned that these proteins spot stalled RNAPII whether or not there is damage to DNA — suggesting that a hitch in the transcription process, not necessarily involving damage to the DNA itself, is the cause of subsequent problems. "It appears that XPG is needed to help CSB recognize the stalled RNAPII and carry out its other functions in TCR," says Cooper. "We think that even if CSB is present, if the cell is missing XPG, the CSB won't work as effectively." This could help explain why mutations in the gene for XPG can cause Cockayne Syndrome even when the gene for CSB is normal. Fixing the machineryThe first step in TCR is the recognition that an RNAPII is stalled. Subsequently one of several things must happen. Most theoretical models have assumed that the stalled RNAPII has to be degraded and moved out of the way so the lesion can be repaired, after which transcription of the gene must start over with a new RNAPII.
Other possibilities are that the original RNAPII is somehow made to start up again, bypassing the lesion, or that the RNAPII is made to back up, digesting some of the messenger RNA it has already made, and have another go at transcription. In these cases, the likelihood of errors in the transcribed RNA and eventually in the resulting protein is increased. What Cooper and her colleagues found, however, was a different, unexpected path. It appears that instead of being removed, the stalled RNAPII can be left in place and remodeled by the protein machinery of transcription-couple repair, so that repairs to the DNA can proceed without the loss of the messenger RNA that has already been formed. The researchers created artificial transcription bubbles of DNA in various sizes, in vitro. In this way they discovered that XPG shows a strong preference for binding to bubbles of just the size typical of transcription — that is, the size of the bubble opened by RNAPII. Identification of this bubble, which does not progress when RNAPII is stalled, together with direct interactions between XPG and the RNAPII itself, seems to trigger the cooperation of XPG and CSB. When RNAPII stalled at damaged nucleotides during transcription carried out in vitro, XPG and CSB together formed a large, stable complex with the polymerase. XPG's function during damage removal is to cut the damage-containing DNA strand on one side of the lesion, as part of a complex cut-and patch process. But when the damage causes stalled transcription this site is inside the RNAPII, which has closed itself around the two strands of DNA with a "clamp module." Indeed, the researchers found that RNAPII bound to a transcription bubble prevented incision by XPG. To their surprise, however, adding another large protein machine called Transcription Factor IIH (TFIIH) allowed XPG to cut the DNA strand at the appropriate place — without removing the RNAPII. The best explanation is that TFIIH causes a conformational change, a "remodeling" of the RNAPII, which lifts the clamp module out of the way. This finding is especially significant because XPB and XPD, the other two proteins associated with xeroderma pigmentosum that can cause Cockayne Syndrome when defective (in addition to XPG), are essential components of TFIIH. TFIIH does not recognize stalled RNAPII on its own. It appears that either CSB or XPG is needed to recruit TFIIH to the stalled polymerase, and it's likely they do this more efficiently when working together. The new findings suggest that one of the major functions of TFIIH — previously unsuspected — is to clear the way for transcription-coupled repair by opening up the RNAPII without destroying it. "This new mechanism means that transcription-coupled repair is much more efficient than other processes for resolving stalled transcription, since it can operate with RNAPII still in place, so that transcription to that point is not lost," says Cooper. "Remodeling would thus be the pathway of choice, with alternative methods available in a pinch. Only if all else fails would the RNAPII be destroyed, forcing transcription to start fresh." In this first in a series of papers on the mechanisms of TCR, Cooper and her colleagues have shed early light not only on how the mysterious process of transcription-coupled repair works at the molecular level, but also on the underlying cause of the transcription defects that accumulate after birth in Cockayne Syndrome, which result in cell death leading to CS's characteristic neurological decline and wasting. Further work may provide insight into the processes of aging itself. "Recognition of RNA polymerase II and transcription bubbles by XPG, CSB, and TFIIH: insights for transcription-coupled repair and Cockayne Syndrome," by Altaf H. Sarker, Susan E. Tsutakawa, Seth Kostek, Cliff Ng, David S. Shin, Marian Peris, Eric Campeau, John A. Tainer, Eva Nogales, and Priscilla K. Cooper, appears in the 28 October 2005 issue of Molecular Cell. Berkeley Lab is a U.S. Department of Energy national laboratory located in Berkeley, California. It conducts unclassified scientific research and is managed by the University of California. Visit our website at http://www.lbl.gov. |
||||||||||||||||
| Top | ||||||||||||||||