The role of epimorphin as a extracellular morphogen was originally identified in studies of lung branching morphogenesis. The same molecule was subsequently identified as syntaxin-2, a member of the syntaxin family of vesicle fusion proteins. Lung organ cultures, which contain both epithelial and mesenchymal cells, can branch in 3D collagen if recombinant growth factors are provided, but this activity is completely blocked by the addition of anti-epimorphin antibodies.
Since that
initial discovery, epimorphin has been found to play a role in morphogenesis
of many tissues, although it is the mammary gland in which the action
of epimorphin has been most clearly defined. In the mammary gland, epimorphin
expression reflects its role in mesenchymal-to-epithelial signaling: in
tissue sections of postnatal mouse mammary glands, epimorphin is expressed
within the stroma and around the epithelial ducts, while staining of isolated
primary cells showed epimorphin expression in fibroblasts and myoepithelial
cells, but not in luminal epithelial. During lactation, epimorphin can
also be found around luminal epithelial cells, in the ductal lumina, and
in the milk. As with lung organ cultures, branching morphogenesis can
be induced in “tissue organoids” isolated from the mammary
gland, if the organoids are cultured in 3D collagen and treated with EGF,
HGF, FGF-1, or KGF; also as with the lung organ cultures, growth-factor
induced branching could be inhibited by the addition of anti-epimorphin
antibodies.
Evaluation of a number of mammary epithelial cell lines showed that a
majority expressed epimorphin, while no epimorphin expression was detectable
in others. However, for both types of cell lines, epimorphin activity
was required for morphogenesis. Growth factor-induced branching of epimorphin-expressing
cell lines could be inhibited with antibodies against epimorphin and the
particular exogenous growth factor used, but not with antibodies against
any other growth factor (i.e., HGF-induced branching could be inhibited
by anti-HGF antibodies, but EGF-induced branching was not blocked by anti-HGF
antibodies). These results demonstrated that epimorphin was not acting
by inducing the expression of alternative growth factors. For non-epimorphin
expressing luminal epithelial cell lines, branching required the addition
of both exogenous epimorphin and any one of several different growth factors.
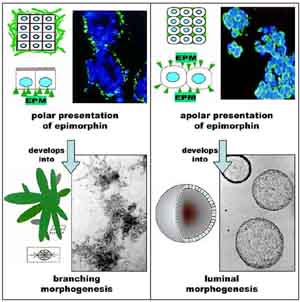
Experiments using recombinant epimorphin also revealed its potential to
mediate dramatically different morphogenic processes depending upon orientation
of presentation. Presented in a polar fashion to the basal surface of
the cell clusters, epimorphin stimulated branching morphogenesis; presented
in an apolar fashion around all surfaces of the cells in the clusters,
epimorphin instead stimulated the formation of cystic structures with
large central lumen (Figure). For both types of morphogenesis, the morphogenic
signal was provided by epimorphin, while the extent of growth (branch
length elongation or increase in luminal diameter) was proportional to
the proliferative activity induced by the particular growth factor.
|
|
|
Recent Publications: Radisky D.C., Hirai Y., Bissell M.J. Delivering the Message: epimorphin and mammary epithelial morphogenesis Trends Cell Biol. 2003 Aug;13(8):426-34. Simian M, Hirai Y, Navre M, Werb Z, Lochter A, Bissell MJ. The interplay of matrix metalloproteinases, morphogens and growth factors is necessary for branching of mammary epithelial cells. Development. 2001 Aug;128(16):3117-31. Hirai Y, Radisky D, Boudreau R, Simian M, Stevens ME, Oka Y, Takebe K, Niwa S, Bissell MJ.Epimorphin mediates mammary luminal morphogenesis through control of C/EBPbeta. J Cell Biol. 2001 May 14;153(4):785-94. |