| April 30, 2004 | science beat | | lab a-z index | lab home |
|
|||
| Asf1: A Protein in the Thick of Things | ||||||||||||||||||||||||||||||||||||||||
| Contact: Paul Preuss, paul_preuss@lbl.gov | ||||||||||||||||||||||||||||||||||||||||
| Picture the crew of a big 19th-century square-rigger in action, with hundreds of sailors swarming over the rigging, setting and trimming the sails, each with a specific task but all working in synchronization to keep the ship safely under way.
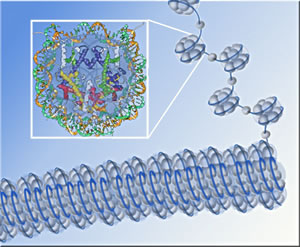
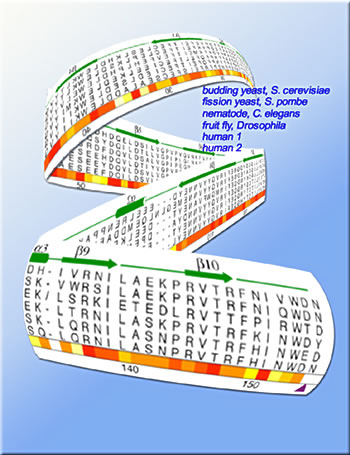
It's not too different from the way a crew of proteins cooperates to keep cells afloat: among other tasks, these gangs of proteins build scaffolding on which DNA winds itself to make chromatin, the stuff of chromosomes; they remodel the chromatin, exposing and opening up double-stranded DNA to make genes available for transcription or sealing DNA strands to silence genes; they man the checkpoints that regulate cell reproduction; and they repair the DNA when it breaks. "One protein that's found at the nexus of many of these processes is a small, compact protein known as Asf1," says Paul Kaufman, a staff scientist in Berkeley Lab's Life Sciences Division and an associate adjunct professor in UC Berkeley's Cell and Molecular Biology Department. The main role of Asf1 ("Anti-silencing factor 1") is as a histone chaperone, helping to deposit histone proteins on DNA strands to form nucleosomes, the protein-DNA units that when linked together make up chromatin. Chromosome assembly begins when eight histone subunits are brought together and a double strand of DNA loops around them twice — more precisely, one and two-thirds times — like thread around a spool. The result is a nucleosome. The continuous DNA strand connects the nucleosomes like beads on a string, and this DNA-protein beaded string is rolled up into a cylindrical rope-like structure, chromatin, which is further folded and looped into the compact mass of the chromosome. Besides helping deposit histones from which nucleosomes are assembled, Asf1 involves itself in other critical functions. First identified in baker's yeast, genes for Asf1 have been identified in many other eukaryotes (organisms whose cell nuclei are enclosed in membranes). In fact, says Kaufman, "all eukaryotes have at least one version of the gene. Some, including humans, have two."
Kaufman's lab joined forces with the lab of James Berger of Berkeley Lab's Physical Biosciences Division, an associate professor of biochemistry and molecular biology at UC Berkeley, to investigate the structure and function of this key protein, in collaboration with Peter Adams from the Fox Chase Cancer Center in Philadelphia and Steven Brill of Rutgers University. "Our first step was to validate the biological importance of Asf1, using standard biological and biochemical means," says Kaufman. "We were particularly interested in isolating the 'core' of this protein, the part that has been conserved during evolution." Kaufman explains that the protein's first 155 amino-acid residues, counting from the exposed amino-group end of the string (the N-terminal), are highly conserved in virtually all organisms. Their 65 to 90 percent identity among organisms separated by hundreds of millions of years of evolution or more is clear evidence of this part of the protein's fundamental importance to cell function. The rest of the sequence (the C-terminal) varies widely among organisms, and in at least one, the parasite Leishmania major, it is missing altogether. "We let evolution tell us where to cut the tails off," Kaufman says, describing how the researchers synthesized the isolated core fragment of Asf1, using recombinant DNA technology. In vitro, the resulting purified Asf1 fragments were able to bind to histones as readily as the complete Asf1 did, and they interacted equally well with a cell-cycle-checkpoint enzyme. Mutant yeast cells that lack Asf1 are sensitive to DNA-damaging agents like methyl methanesulfonate and hydroxyurea. But when the researchers introduced a plasmid containing a modified gene for encoding the Asf1 fragment into the mutant cells, the Asf1 core alone restored resistance to damaging agents equally as well as the entire protein. The core of Asf1 also stimulated another histone deposition protein, CAF-I, which plays an important role in chromatin assembly during DNA replication. Together, these findings confirmed that the core of Asf1 was definitely the business end of the protein.
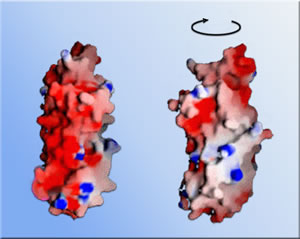
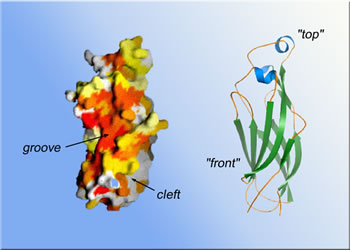
Once these and other functions of Asf1 and its core unit had been established beyond doubt, the researchers set out to solve the core's structure, seeking to understand how it performs its multiple tasks. Sally Daganzo, a member of Kaufman's lab now at the University of California at San Francisco, got help from Berger's lab in growing crystals of the Asf1 core regions and introducing the heavy atoms needed for x-ray crystallography. "By using beamline 8.3.1 at the Advanced Light Source, we were able to get the data extremely rapidly," says Kaufman. The result was a 3-D model of the Asf1 core at a resolution of 1.5 angstroms
(one and a half ten-billionths of a meter). "We identified conserved
salient features on the surface of the model, looking for clusters of
charged or hydrophobic amino-acid residues, anything that looked like
a docking site," Kaufman says. Three regions stood out: a hydrophobic
("water-fearing") concave groove on the arbitrary front of the
molecule; a nearby area of strong negative electric charge; and, on the
opposite side of the groove, a deep pocket lined with hydrophobic residues
where the ends of the core's chain of amino-acid residues come together
at its bottom.
"In the case of Hir and HIRA, Asf1 may act as a histone-reloading protein," Kaufman says. "Hir and HIRA continually deposit histones as they are building chromatin. Asf1 may be the carrier that reloads their magazines." He emphasizes that "the full impact of what we know hasn't come out yet. We are in the process of looking carefully into how this one protein manages to find itself a central player in so many different processes." Additional information
|
||||||||||||||||||||||||||||||||||||||||
| Top | ||||||||||||||||||||||||||||||||||||||||