| July 26, 2006 | science@berkeley lab | | lab a-z index | lab home |
|
|||
| Secrets of a Cellular Machine New Clues to the Architecture of Flagella and Cilia |
|||||||||||||||||||||||||||||||||||
| Contact: Paul Preuss, paul_preuss@lbl.gov | |||||||||||||||||||||||||||||||||||
Kenneth Downing and Haixin Sui of Berkeley Lab's Life Sciences Division have pioneered the use of cryo-electron tomography to examine the ubiquitous protein structures called axonemes, which form the cores of the cilia and flagella of eukaryotic cells.
Axonemes are some of nature's largest molecular machines. Their principal structural elements are microtubules, tough and versatile protein assemblies that perform many cellular roles, notably as major components of the cell skeleton. In 1998 Downing and Eva Nogales, then a scientist in his group, with colleague Sharon Wolf, first revealed the structure of alpha and beta tubulins, the protein dimers from which microtubules are constructed. In 2002 Downing and Huilin Li, also a scientist in his group, published details of a microtubule's structure at eight-angstrom resolution, better than twice that ever obtained before. "In the present work Haixin Sui and I were initially looking to follow up the earlier work on tubulin," Downing says. "In mammals tubulin comes in many forms, so we intended to isolate the simple form in sea urchin eggs in hopes of making better crystals. It turned out that we also collected a lot of sea urchin sperm, which are an excellent source of axonemes." Whips and eyelashesLacking legs or flippers, many single-celled eukaryotes (eukaryotic cells are those with nuclei) get around using flagella and cilia, Latin for "small whips" and "eyelashes." Nor could complex creatures, including human beings, survive without these powerful molecular machines. The cilia that sprout thickly from cells that line the lungs and other organs wave as rhythmically as sea grass in the tide to dislodge and sweep away litter. Flagella thrash energetically to propel sperm. Except for length and number per cell, flagella and cilia are similar and share a common structure. At the center of each is the axoneme, a tough, flexible bundle of microtubules encased by a membrane. Other proteins connect the microtubules in the axoneme together or move over them, causing them to bend and slide against each other in a rhythmic beating motion. "The basic axoneme plan has been known for forty years, from biochemistry and low-resolution electron microscopy," says Sui. "But finding out which proteins are located where, and even learning the identities of many of the proteins, has long frustrated researchers." "Resolution was the challenge," Downing says. "Conventional electron microscopy just couldn't see the details." The latest high-resolution results with cryo-electron tomography offer new insights and promise new understanding of these vital cellular structures.
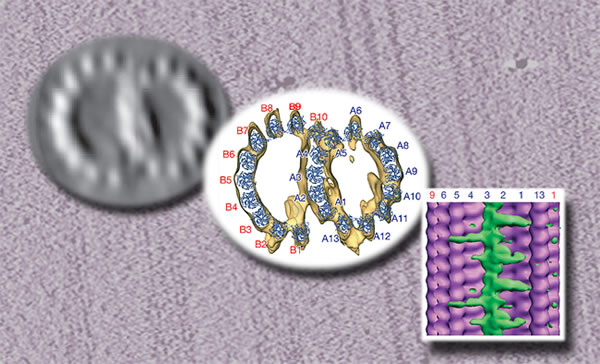
Single microtubules consist of tubulin dimers strung in what are called protofilaments, usually thirteen parallel protofilaments arranged to form an openwork cylinder. An axoneme has two such singlet microtubules in the center, ringed by nine doublet microtubules, which are basically two singlets attached along their length. The whole coaxial bundle is linked together and activated by other proteins. (Microtubules come in triplets too. The structure that attaches the axoneme to the cell, called the basal body, consists of nine relatively short triplets. A similar structure, the centriole, organizes the microtubules that attach to chromosomes during cell division.) "Given that we had a large supply of axonemes in the form of sperm from local sea urchins, it was a logical extension of the microtubule singlet work previously done in this lab to study microtubule doublets," says Sui. The Downing laboratory has been a pioneer in the use of cryo-electron crystallography, in which protein structures are reconstructed in silico using data from many images and diffraction patterns of tiny crystals of the protein. They also use single-particle methods, in which structures are visualized using images of hundreds of "particles," individual proteins frozen in random orientations on a flat surface. Says Downing, "Haixin persuaded me that we could use our new electron microscope to do high-resolution cryo-electron tomography on these structures." Cryo-electron tomography goes a step farther than single-particle reconstruction, starting with one or a few high-quality samples (typically complex protein structures) and tilting these under the microscope's electron beam to obtain the data for computer reconstruction of a three-dimensional image. The computer can also combine, or "dock," models constructed using several different methods, to achieve higher resolution in some parts of a structure and to provide clues to the identity of unknown structures. Homing in on doublet structureSui first prepared the sea urchin doublets for electron tomography by separating the axonemes from the sperm heads and using solutions of salts and sugars to remove the axoneme membrane. The process also removed many associated proteins whose identity was already known, including the molecular motors kinesin and dynein. But along with the skeletal microtubules, other key proteins remained. Using the tomographs, the researchers were able to pinpoint the spacing and arrangement of the overlapping microtubules that form the doublet. Viewed in cross section from the top, both are slightly squeezed. One of the two, the A tubule, corresponds to a complete singlet having 13 protofilaments; the other, the B tubule, would measure 15 protofilaments around if it were complete, but five of these are lacking where the two tubules are joined at the partition. The partition between the tubules is formed by four protofilaments of tubule A, three of which form a stable ribbon, a sort of backbone for the doublet. At one end of the partition there's an opening between the tubules — here, researchers working from lower-resolution micrographs had long assumed another protofilament must lie.
"We showed there is no protofilament there," Downing says. "Instead, there's a linker protein that crosses the gap between the tubules. We also have hard evidence for the identity of some of the inner binding proteins," which are concentrated along the sides of the inner partition. These binding proteins add stiffness to the partition, which causes the two tubules to appear squeezed or flattened, profoundly limiting the way the doublet can bend. The cryo-electron tomogram yields a three-dimensional model of the doublet that can be rotated and viewed from any angle; seen from the side, the partition shows protein structures repeating in multiples of eight nanometers, the length of the tubulin dimers that make up the protofilaments. Some processes bridge from the backbone ribbon to protofilaments on either side; the shorter ones repeat every eight nanometers (corresponding to a tubulin dimer) and the longer ones repeat every 16 nanometers (two dimers). There are several other protein groups inside the wall of the A tubule. Evidence from transmission electron microscopy suggests that some of the proteins inside the A tubules are various forms of tektin. Biochemical evidence and immunofluorescence microscopy indicate that the linkers, which extend outward from the partition to close the gap between tubules A and B (and also repeat at 16-nanometer intervals), are probably made up of a pair of polypeptides, known in sea urchins as Sp77 and Sp83.
"This is the clearest anyone has ever seen inside the doublet," says Downing, "and it gives us fundamental structural information, answering a number of questions about axonemes. Side by side doublets slide up and down relative to each other, but A and B tubules within the doublets do not slide against each other, as some have proposed. The stiffness of the tubules determines the plane in which the doublet can bend, and the linker protein across the gap between the tubules allows even an individual doublet to twist. Because of their arrangement in the axoneme, the axoneme as a whole twists under stress." These first results with cryo-electron tomography open many paths to knowledge about the vital axoneme structure. "We already have hints about the structure of linkages between the doublets in the axoneme," says Sui, "which may give us information about the coordinated sliding of the doublets." Says Downing, "The axoneme is a basic structure in all eukaryotes. Our eventual goal is to find out how it evolved and why it has been conserved since the beginning." Additional information
|
|||||||||||||||||||||||||||||||||||
| Top | |||||||||||||||||||||||||||||||||||