| November 3, 2005 | science@berkeley lab | | lab a-z index | lab home |
|
|||
| Ratcheting, Swiveling, Opening the Groove Highest Resolution Yet of the Intact Ribosome Reveals Machinery of the Protein Assembly Line |
||||||||||||||||||||||||||||||||||||||||
| Contact: Paul Preuss, paul_preuss@lbl.gov | ||||||||||||||||||||||||||||||||||||||||
| Proteins are encoded in the genomes of every living thing, but how is this information translated into protein structures? Cellular organelles called ribosomes do the work; despite some differences, the ribosomes of everything from single-celled bacteria to multicellular animals like us are remarkably similar.
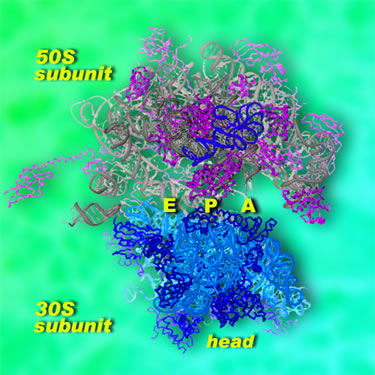
Ribosomes are unusual cellular machines in that they are not made mostly of proteins; instead, two-thirds of the ribosome consists of a special form of RNA. Detailed structures of the ribosome's two main components have been created in recent years, but these are a little like having the plans for two halves of a car without knowing how they fit together. "People have been working on protein synthesis for many decades," says Jamie Cate, a staff scientist in Berkeley Lab's Physical Biosciences Division and an assistant professor of biochemistry, molecular biology, and chemistry at the University of California at Berkeley. "The problem boils down to determining the structure of the intact ribosome at a high enough resolution to see exactly what's going on." Now, through x-ray crystallography of Escherichia coli ribosomes in two distinct configurations, Cate and his colleagues have determined their structures to a resolution of 3.5 angstroms, the highest-resolution structure of the intact ribosome ever achieved. (An angstrom is a ten-billionth of a meter, about the radius of the smallest atoms, or just smaller than chemical bonds.) "This is the first functional atomic-resolution model of the intact ribosome that we can refine and validate," says Cate. "Comparing our structures with each other and with other models allows us to predict how the ribosome translates chemical energy into mechanical motion." Modeling the E. coli ribosome in two configurations provides snapshots of the machinery by which the messenger RNA (mRNA), which brings the genetic information from the cell's DNA, and the transfer RNAs (tRNAs), which bring individual amino acids to be assembled into a protein, are moved together through the ribosomal assembly line. Looking from the outside inThe ribosome consists of two subunits, large and small, which remain apart until they join to work together during protein synthesis. In bacteria the small subunit is designated 30S and the large subunit 50S; the intact ribosome is labeled 70S. (S refers not to a unit's dimensions but to its sedimentation rate in a centrifuge, thus its mass.) High-resolution structures have previously been determined for the individual subunits, but not for the intact state in which protein synthesis occurs.
Messenger RNA is produced by the cell's DNA in a form coded like a tape, in which each codon, or group of three nucleotides, represents one amino acid. When the 30S subunit of a ribosome encounters a length of mRNA, it moves along it until it finds the codon that means "start." To start a protein, one end of an "initiator" tRNA binds to the small 30S subunit at a site called P (for peptidyl or "peptide-forming"), situated near the mRNA. The other end of the initiator tRNA carries an amino acid that is always the first in the peptide chain — in bacteria, a special form of methionine. (Proteins are built from just 20 amino acids, with one or more tRNAs specific to each.) The larger 50S subunit of the ribosome then joins the 30S subunit, and they sandwich the tRNA between them. Fed in by an elongation factor, a new tRNA binds to a site labeled A (for aminoacyl or "amino acid") on the small subunit. A peptide bond forms between the first and second amino acids, and then the ribosome moves the assembly line forward by one codon position; the initiator tRNA moves to site E (for "exit") and the next tRNA moves from site A to site P. As the ribosome moves along, reading the mRNA, the cycle repeats, shuttling tRNAs from site A to site P to site E — from the amino-acid site to the peptide-forming site to the exit ‑‑ while the protein elongates one amino acid at a time. The whole process is powered by the chemical fuel GTP, carried by the elongation factors attached to the tRNAs. Translocation and elongation require stable and highly flexible interactions between the ribosome's two subunits, which work together until the small subunit finally encounters the "stop" codon on the mRNA. This signal summons a set of proteins that release the newly completed protein and break apart the 30S and 50S ribosomal subunit so that they can go their separate ways. Inside the machineTo learn how the subunits work together during translocation, Cate and his colleagues first had to make thousands of extremely high-quality crystals of the intact ribosome, suitable for x-ray crystallography. Crystals consist of repeating unit cells, but in a ribosome crystal each unit cell consists of smaller asymmetric units containing two intact ribosomes — some 100 billion ribosomes in all — "which sounds like a lot," Cate says, "but is actually a fairly low number for one of these large-molecule projects." To his delight, in the 17 identical crystals used for the present work, the twin ribosomes in each asymmetric unit proved to be in strikingly different conformations. "Before coming to Berkeley Lab and UC Berkeley, I spent two years working with crystals that didn't diffract well; the ribosomes are more in control of the crystal than we are. But these were excellent." Cate and his colleagues first assembled models from earlier structures of the ribosome at a resolution of 9 angstroms, which helped orient the two configurations of the intact ribosome. Using beams of hard x-rays, first from the Advanced Light Source's crystallography beamline 8.3.1 and later, for more detailed work, from beamline 12.3.1, named SIBYLS (for Structurally Integrated Beamline for Life Sciences), Cate's team measured the data needed to refine the structure to 3.5-angstrom resolution.
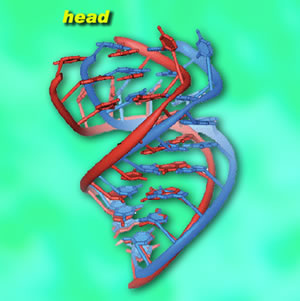
At this resolution individual components were revealed in startling detail, down to individual nucleotides in helical strands of RNA. The small subunit is particularly complex in structure, with features called the head, neck, shoulder, body, platform, and spur. The head is notably active during translocation. Cate's team discovered three helical "tension springs," made of simple RNA helices. They think that one, attached to the neck, is relatively stiff, while the others are flexible. These guide the head as it helps move mRNA and tRNA through the mechanism. The researchers also found that the gap between the small and large subunits is filled with salt water. Since the ribosome's own RNA is infused with magnesium ions and is highly charged, the salt water may act as a kind of "lubricant," locally neutralizing the charge between the subunits to allow them to move in relation to each other and allow tRNAs to slide through the gap. Stations on the assembly lineThe researchers compared their two high-resolution snapshots of the intact E. coli ribosome with a wide range of conformations of other ribosomes; these included lower-resolution x-ray crystallography of Thermus thermophilus and E. coli ribosomes, plus electron microscopy of E. coli, yeast, and mammalian ribosomes. These yielded what Cate calls "global snapshots" and allowed him and his colleagues to deduce how individual parts of the ribosome function during the translocation process.
What emerged from this process is a series like a filmstrip, each frame showing one of the distinct motions of the small subunit head during translocation and elongation. Once the A‑site tRNA attaches to the growing peptide chain, as peptide bond formation occurs, the small subunit ratchets backwards with respect to the large subunit, preparing to move the assembly line of mRNA and tRNAs forward by one codon. Now the head swivels, opening a groove between the P and E sites that "unlocks" the tRNAs and mRNA and allows them to move forward. At this point, the small subunit reverses its motions, resets, and is ready to bind the next tRNA. While this picture of translocation — ratcheting, swiveling, opening the groove, then reversing these three steps — is a far more detailed analysis of protein synthesis than has been possible before, Cate stresses that much remains to be learned. "We would really like to be able to see the bound elongation factors as they move the tRNAs through the ribosome," he says. "We would like to image the proteins that hydrolyze GTP to help move the tRNAs and mRNA. And we would like to examine the mRNA-decoding step — we've shed a lot of light on translocation, but not as much on mRNA decoding." To achieve these goals, these components will have to be co-crystallized, catching complex structures in the act. There is a mismatch in time scale that makes the work inherently challenging, Cate says. "Each step in making a protein takes the ribosome maybe a few milliseconds. Growing good crystals of each of these steps takes days or even weeks." Capturing complexes of ribosomes, mRNAs and tRNAs, elongation factors, and other molecules together in one crystalline form will require techniques for markedly slowing these lightning-fast reactions. The power and convenience of the ALS beamlines for crystallography are what encouraged Cate to move his research to Berkeley from the East Coast. The National Institutes of Health, through the National Cancer Institute, and DOE funded Cate's work on the 8.3.1 and SIBYLS beamlines. Initial construction of the SIBYLS beamline was also funded by DOE. "Structures of the bacterial ribosome at 3.5 Å resolution," by Barbara S. Schuwirth, Maria A. Borovinskaya, Cathy W. Hau, Wen Zhang, Antón Vila-Sanjurjo, James M. Holton, and Jamie H. Doudna Cate, appears in the 4 November issue of Science. Additional information
|
||||||||||||||||||||||||||||||||||||||||
| Top | ||||||||||||||||||||||||||||||||||||||||