|
January 29, 2003
|
||
|
 |
 |
|
What a difference development makes How a single gene makes different forms of protein as needed |
||
| Contact: Paul Preuss, paul_preuss@lbl.gov | ||
|
|
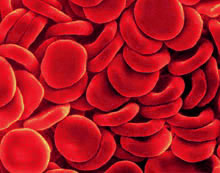
To deliver oxygen throughout the body, red blood cells must repeatedly squeeze through the narrow passages of tiny blood vessels, then rapidly recover their shape; if their membranes couldn't stand up to mechanical stress, they would fragment easily. What makes a red cell's membrane tough and flexible are "skeletal" proteins like actin and spectrin. To form effective linkages between these two kinds of protein, a third protein is needed, known as protein 4.1R. In the stem-cell precursors of red cells, which are found in the bone marrow, actin-spectrin linkages are not needed and might interfere with the process of development. Remarkably, these early differentiating cells produce a different form of 4.1R, one whose spectrin-actin binding domain doesn't work. John Conboy of Berkeley Lab's Life Sciences Division and his colleagues have recently found the mechanism that switches on and off a part of the 4.1R gene that codes for the protein's spectrin-actin binding domain. They have pinpointed a series of interactions — among sequences in the gene with proteins in the cellular environment — that determine what form of 4.1R the gene will express at different stages of the cell's development. (See sidebar, "How red blood cells develop.") Splicing and alternate splicing Conboy's interest in protein 4.1R began years ago when he was a postdoctoral fellow in Y. W. Kan's lab at the University of California at San Francisco, UC's leading medical campus. In collaboration with biophysicist Mohandas Narla, he initiated studies of the protein 4.1R gene in order to understand a rare form of hemolytic anemia (in which red blood cells are attacked by the body's own immune system), characterized by unstable red blood cell membranes. "The gene we were studying turned out to be much larger than the average human gene," Conboy says, "extending over about 240 kilobases" — some 240 thousand bases of DNA. "The gene also exhibits an extraordinary degree of complexity, with many alternate splices." Almost all genes require splicing, because in addition to the DNA sequences called exons, which code for a protein's amino acids, they include noncoding sequences called introns. To form the messenger RNA (mRNA) that specifies the order of amino acid residues in the finished protein, intron sequences have to be clipped out and coding sequences spliced together. Many genes are hard-wired to express a single mRNA product, in which all of the exons are obligatorily spliced together. By contrast, other genes can generate multiple mRNAs by alternative splicing, which occurs when one or more selected exons are conditionally spliced in or clipped out of the mRNA. At present it's estimated that at least half of the roughly 30,000 genes in the human genome are subject to alternative splicing. In many cases alternative splicing is crucial to the development, health, and survival of the organism. Conboy has concentrated on alternate splicing during tissue differentiation. "When a tissue is differentiating, many changes in gene activity are occurring. Many researchers have focused on which genes are being turned on and off, but alternative splicing in individual genes may be just as important," he says. The versatile 4.1R gene The 4.1 family of proteins is found in many different species and is important in the cellular structures of many different kinds of cells, including the mechanisms of chromosomal organization in nucleated cells. But the best-known member of the family is protein 4.1R, which acts as an anchor for the skeletal proteins of the red cell membrane.
The gene for protein 4.1R has some two dozen exons, many of which are subject to alternative splicing. Conboy and his colleagues have zeroed in on one of these, labeled exon 16, which is suppressed while the red cell is developing but must be switched on in the mature cell. He says, "The result of a failure to make this switch is an unstable red cell membrane." The first step in translating the information encoded in a gene into a working protein is to assemble a strand of messenger RNA that precisely specifies the protein's amino-acid sequence. Precursor mRNAs (pre-mRNAs), raw copies of the gene that contain both the coding exons and noncoding intron sequences, must be edited and assembled by a complex of proteins and RNA called the spliceosome — what Conboy calls "a big protein machine" — inside the cell nucleus. Exon 16 in the 4.1R gene contains a number of nucleotide sequences that are very similar among different vertebrate species, including human, mouse, cow, frog, and even chicken. In the language of genetics, these sequences are highly conserved, suggesting that they play a persistent and fundamental role that emerged early in evolution.
Much of exon 16 consists of an extraordinarily well conserved sequence designated CE16. Approximately 40 nucleotides long, CE16 has a special function: it is a "splicing silencer element" that, when activated, prevents the exon from being recognized by the spliceosome and spliced into the messenger RNA. A 4.1R protein specified by mRNA that lacks the exon-16 sequence cannot bind spectrin and actin in the red cell membrane. When CE16 is turned off, however, exon 16 is recognized and its information is spliced into mRNA; the resulting 4.1R protein has a functioning binding domain, essential for constructing the tough, flexible membranes of mature red blood cells. Minigenes and mutations pinpoint a protein Conboy and his colleagues devised a series of experiments that not only identified CE16 as the crucial splicing silencer but also pinpointed the factors that trigger it during red cell differentiation. Instead of working with the entire large 4.1R gene, the researchers constructed a "minigene" with only three exons, including exon 16. They manipulated various nucleotide sequences in the minigene and the pre-mRNA it produced and observed the results in human and other cells grown in vitro. In some mutations, the researchers replaced virtually the entire CE16 silencer region with a neutral sequence (borrowed from the fruit-fly genome) of equal length. With the original CE16 sequence missing, the mutated exon was expressed efficiently. This confirmed that CE16 is needed to prevent the splicing of the exon into mRNA. In other mutations just four or five nucleotides were changed at specific sites. These mutations homed in on what turns the silencer on or off. The researchers found that the mutations which destroyed silencing activity had caused modifications of the pre-mRNA in regions where a specific protein binds during cell differentiation. The protein involved is called heterogeneous nuclear ribonucleoprotein A1 (hnRNP A1). HnRNPs cooperate with the spliceosome, the molecular machinery that clips out introns and splices together the exons to form mRNA; they bind to the raw pre-mRNA transcripts of the gene and are active in RNA splicing. With additional experiments the researchers confirmed that hnRNP A1 binds directly to the CE16 silencer and that this interaction prevents splicing of exon 16. Levels of the A1 protein are high during the early stages of red blood cell differentiation, but they decrease dramatically as the cells mature. The effect of decreasing hnRNP A1 is to inactivate the CE 16 silencer, allowing exon 16 splicing to occur. In turn this allows synthesis of 4.1R proteins with functional spectrin-actin binding domains — just when they are needed to produce cells with tough, flexible membranes. Conboy and his colleagues showed that the mechanism of this critical step in blood-cell development is essentially identical among mammals, birds, and amphibians.
These studies reinforce the notion that the DNA sequences in a gene's exons play multiple roles in biology. "The most obvious role for DNA sequences in the past was that they specify the amino-acid sequences in the proteins they encode. Now we know that nucleotide sequences also play an important role in regulating the decision to splice the exons in which they reside," Conboy says. "The CE16 sequence in exon 16 of the 4.1R gene is an excellent example: it encodes a critical portion of the protein domain required for binding spectrin and actin, and also helps to regulate exon 16 splicing." For recent work on the 4.1R gene, Conboy emphasizes the contributions of his colleagues Victor Hou, now at Children's Hospital Oakland, and the others in the splicing research team at Berkeley Lab. Akila Mayeda of the University of Miami School of Medicine and Adrian Krainer of Cold Spring Harbor shared their expertise on alternative splicing. Mark Koury of the Department of Medicine, Vanderbilt University, provided red blood cells precursors at different stages of development. "Decrease in hnRNP A/B expression during erythropoiesis mediates a pre-mRNA splicing switch," by Victor C. Hou, Robert Lersch, Sherry L. Gee, Julie L. Ponthier, Annie J. Lo, Michael Wu, Chris W. Turck, Mark Koury, Adrian R. Krainer, Akila Mayeda, and John G. Conboy, appears in the November 15, 2002, issue of the EMBO Journal. Additional information |
||||||||||||||||||||||||||||||||||||||